Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in collaboration with the World Health Organization, GA(2)LEN and AllerGen)
1
2008
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
Prevalence of self-reported allergic rhinitis in eleven major cities in china
1
2009
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
The link between allergic rhinitis and allergic asthma: a prospective population-based study. The copenhagen allergy study
1
2002
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
Allergic rhinitis and asthma comorbidity in a survey of young adults in Italy
1
2005
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
Epidemiologic evidence for asthma and rhinitis comorbidity
1
2000
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
The alliance of genes and environment in asthma and allergy
1
1999
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
Evidence for common genetic elements in allergic disease
1
2000
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
111(2 Suppl): 11S-16S
1
1997
... 变应性鼻炎(allergic rhinitis,AR)是由IgE介导的I型变态反应引起的鼻部炎性疾病,主要症状为鼻痒、流涕、喷嚏和鼻塞[1].该病目前在全球的流行明显增加,在我国患病率为8.7%~24.1%[2].同时,许多流行病学研究发现鼻炎常伴随着哮喘的发生发展[3,4,5].AR和哮喘均为受多因素影响、基因与环境共同作用的变态反应疾病[6],虽然AR与哮喘的变态反应症状分别表现在上、下呼吸道,但两者不仅遗传背景相似[7],并且呼吸道黏膜结构及功能相似,因此提出了“同一气道,同一疾病”的概念[8]. ...
Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines
1
2002
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
Cloning and characterization of a sub-family of human toll-like receptors: hTLR7, hTLR8 and hTLR9
1
2000
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
A synthetic Toll-like receptor 2 ligand decreases allergic immune responses in a mouse rhinitis model sensitized to mite allergen
2
2008
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
TLR2 agonist ameliorates established allergic airway inflammation by promoting Th1 response and not via regulatory T cells
2
2005
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Innate immune response of alveolar macrophage to house dust mite allergen is mediated through TLR2/-4 co-activation
2
2013
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Human CD4+ T cells express TLR5 and its ligand flagellin enhances the suppressive capacity and expression of FOXP3 in CD4+CD25+ T regulatory cells
1
2005
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
PMID: 16225766
3
2004
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... [15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR通过多种识别分子与其配体结合,经多种衔接分子将信号转导至细胞内,如IL-1受体相关激酶(IL-1 receptor-associated kinase,IRAK)、肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor,TRAF6)等,激活NF-κB或丝裂原活化蛋白激酶等,诱导免疫相关基因的表达[35].TLR通路中的细胞因子在信号传导中发挥重要作用.细胞内TLR2/4信号传导起始的关键分子机制是TLR2/4、TIRAP和MyD88三者的TIR结构域的特异性组装[19].CD14与TLR2、TLR4通过相互间的协同作用发挥其生物学功能[15,22]. ...
Toll-like receptor 7-triggered immune response in the lung mediates acute and long-lasting suppression of experimental asthma
2
2010
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
CpG oligodeoxynucleotides can reverse Th2-associated allergic airway responses and alter the B7.1/B7.2 expression in a murine model of asthma
2
2000
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
Allergen-independent immunostimulatory sequence oligodeoxynucleotide therapy attenuates experimental allergic rhinitis
2
2004
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
Toll/interleukin-1 receptor (TIR) domain-mediated cellular signaling pathways
3
2015
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR通过多种识别分子与其配体结合,经多种衔接分子将信号转导至细胞内,如IL-1受体相关激酶(IL-1 receptor-associated kinase,IRAK)、肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor,TRAF6)等,激活NF-κB或丝裂原活化蛋白激酶等,诱导免疫相关基因的表达[35].TLR通路中的细胞因子在信号传导中发挥重要作用.细胞内TLR2/4信号传导起始的关键分子机制是TLR2/4、TIRAP和MyD88三者的TIR结构域的特异性组装[19].CD14与TLR2、TLR4通过相互间的协同作用发挥其生物学功能[15,22]. ...
FcepsilonR1 and toll-like receptors mediate synergistic signals to markedly augment production of inflammatory cytokines in murine mast cells
2
2006
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,20,21,22]等在信号传导中发挥十分重要的作用. ...
Regulation of allergic airway inflammation through Toll-like receptor 4-mediated modification of mast cell function
2
2006
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... ,21,22]等在信号传导中发挥十分重要的作用. ...
Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2
2
2003
... 研究发现,Toll样受体(Toll-like receptors,TLR)及其通路与鼻部炎性疾病的发生发展相关[9,10].TLR能通过调控多种与AR发生密切相关的细胞如树突状细胞、肥大细胞和调节性T细胞(regulatory T-cells, Treg),参与Th1/Th2免疫反应平衡的调控.研究显示[11,12,13,14,15,16,17,18]TLR2、TLR5、TLR7、TLR9对变应性炎症主要起负性调节作用,通过诱导Treg细胞扩增等机制,促使免疫系统向Th1分化,抑制或逆转Th2反应.TLR通路中的一些细胞因子比如MyD88[19]、TIRAP[20,21]、CD14[15,16,17,18,19,20,21,22]等在信号传导中发挥十分重要的作用. ...
... TLR通过多种识别分子与其配体结合,经多种衔接分子将信号转导至细胞内,如IL-1受体相关激酶(IL-1 receptor-associated kinase,IRAK)、肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor,TRAF6)等,激活NF-κB或丝裂原活化蛋白激酶等,诱导免疫相关基因的表达[35].TLR通路中的细胞因子在信号传导中发挥重要作用.细胞内TLR2/4信号传导起始的关键分子机制是TLR2/4、TIRAP和MyD88三者的TIR结构域的特异性组装[19].CD14与TLR2、TLR4通过相互间的协同作用发挥其生物学功能[15,22]. ...
e1-8
1
2008
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Polymorphisms in the toll-like receptor 2 subfamily and risk of asthma: a case-control analysis in a Chinese population
3
2010
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
... [24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
An association study of asthma and total serum immunoglobin E levels for Toll-like receptor polymorphisms in a Japanese population
1
2004
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Association analysis identifies TLR7 and TLR8 as novel risk genes in asthma and related disorders
1
2008
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
CD14 and toll-like receptor gene polymorphisms, country living, and asthma in adults
1
2009
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Polymorphisms in interleukin-1 receptor-associated kinase 4 are associated with total serum IgE
1
2009
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Association of IL33-IL-1 receptor-like 1 (IL1RL1) pathway polymorphisms with wheezing phenotypes and asthma in childhood
1
2014
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Identification of polymorphisms in the Toll-like receptor gene and the association with allergic rhinitis
2
2010
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Association of polymorphisms of toll-like receptor 4 with a reduced prevalence of hay fever and atopy
1
2008
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Toll-like receptor gene polymorphisms are associated with allergic rhinitis: a case control study
1
2012
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
PMID: 18360565
1
2005
... 目前已有多项在不同人群中的研究证实TLR及其通路基因多态性(single nucleotide polymorphisms, SNPs)与哮喘的相关性[23,24,25,26,27,28,29],然而与AR的关联研究报道较少[24,30,31,32,33].本研究选择TLR通路中与变态反应疾病密切相关的TLR2、CD14作为候选基因,筛选6个SNP位点,探讨与AR的相关性及与鼻炎相关临床表型(包括鼻炎严重度、伴随哮喘、血清总IgE及特异性IgE水平等)的相关性. ...
Visual analog scales can assess the severity of rhinitis graded according to ARIA guidelines
1
2007
... 1.2 标本收集和DNA提取 对病例及对照者进行问卷调查,问卷的主要内容包括一般情况、人口学特征、疾病的症状、体征、病史、家族史及伴随疾病.视觉模拟评分(VAS):从0 cm(无影响)至10 cm(影响最重)评价患者对鼻部症状的主观感觉,包括流涕、喷嚏、鼻塞、鼻眼痒及总的鼻部症状.VAS评分<5诊断为轻度鼻炎,VAS评分≥5为中重度鼻炎[34]. ...
Toll-like receptors: function and roles in lung disease
1
2004
... TLR通过多种识别分子与其配体结合,经多种衔接分子将信号转导至细胞内,如IL-1受体相关激酶(IL-1 receptor-associated kinase,IRAK)、肿瘤坏死因子受体相关因子6(tumor necrosis factor receptor-associated factor,TRAF6)等,激活NF-κB或丝裂原活化蛋白激酶等,诱导免疫相关基因的表达[35].TLR通路中的细胞因子在信号传导中发挥重要作用.细胞内TLR2/4信号传导起始的关键分子机制是TLR2/4、TIRAP和MyD88三者的TIR结构域的特异性组装[19].CD14与TLR2、TLR4通过相互间的协同作用发挥其生物学功能[15,22]. ...
Toll-like receptor 4 or 2 agonists decrease allergic inflammation
1
2005
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Cutting edge: activation of Toll-like receptor 2 induces a Th2 immune response and promotes experimental asthma
1
2004
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
e2
1
2013
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
CD14 and toll-like receptor gene polymorphisms, country living, and asthma in adults
3
2009
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
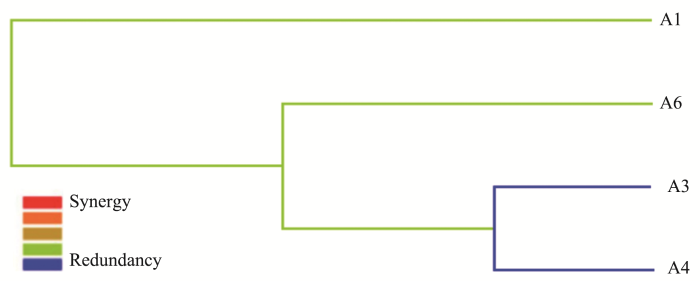
... 本研究采用MDR模型分析发现TLR2与CD14可能存在基因交互作用.已有多项研究发现TLR2与CD14的协同作用,TLR2是LPS的信号转导分子,它的活化有赖于LPS结合蛋白(LPS binding protein,LBP)和CD14的存在.LPS能够诱导肥大细胞释放 TNF-α、IL-5、IL-10和IL-13,激活TLR2或TLR4信号转导通路可以促进Th2型免疫反应,同时也依赖CD14的存在[56].此外有研究显示过敏性疾病中CD14与环境之间同样有交互作用,皮肤点刺阳性患者中携带CD14 rs2569190 CC基因型且在农村环境可降低哮喘发病率[39].通过MDR模型研究,在菲律宾过敏人群[57](CD14与IL4、FCER1B、 IL4RA、 ADRB2)和韩国过敏儿童人群[58](CD14与IL-4Rα、 IL-13、IL-13Rα1、CTLA4)均发现了CD14与过敏相关基因的交互作用.过敏性疾病本身即是基因与环境共同作用,因此基因与基因、基因与环境交互作用将会更全面阐释过敏疾病发生与发展. ...
Atopy and new-onset asthma in young Danish farmers and CD14, TLR2, and TLR4 genetic polymorphisms: a nested case-control study
1
2007
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Toll-like receptors and CD14 genes polymorphisms and susceptibility to asthma in Tunisian children
1
2008
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Toll-like receptor 2 and Toll-like receptor 4 polymorphisms and susceptibility to asthma and allergic rhinitis: a case-control analysis
1
2012
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
A case-control study of innate immunity pathway gene polymorphisms in Puerto Ricans reveals association of toll-like receptor 2?+596 variant with asthma
1
2016
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
Association of single-nucleotide polymorphisms in toll-like receptor 2 gene with asthma susceptibility: a meta-analysis
2
2017
... TLR2在单核-巨噬细胞、内皮细胞、上皮细胞、成纤维细胞等有表达.TLR2识别谱较广, 能够识别细菌肽聚糖、脂蛋白、病毒包膜蛋白等微生物成分并激活信号转导途径,最终导致获得性免疫的发生.大量的研究表明, TLR2对变应性反应有抑制作用[11,12,13,36],但也有报道TLR2对变应性反应有促进作用[37].本研究中rs7656411位点 T等位基因与AR发病存在相关(P=0.035),该位点位于3′侧翼区,可能参与mRNA的转录从而影响基因的表达.目前尚未有rs7656411位点与AR相关性文献报道,但该位点被证实与中国人群哮喘发病相关[24].TLR2的rs4696480位点在加拿大人群中与AR发病无相关[38],597 T/C和1350 T/C 位点在韩国人群中与AR发病无相关,仅单倍型(597C-1350C)有阳性相关[30].TLR2的多态性位点在哮喘病例对照研究中较多,rs13150331、rs2289318、rs3804099位点在法国人群中与哮喘发病相关[39],也有阴性报道[40,41,42,43].最近一篇Meta分析统计TLR2的4个位点rs5743708、 rs3804099、rs3804100和rs4696480对哮喘易感性,发现仅rs4696480位点与哮喘发病相关[44]. ...
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Association between CD14 polymorphism (-1145G/A) and childhood bronchial asthma
2
2016
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Promoter polymorphisms of the CD14 gene are associated with atopy in Pakistani adults
3
2011
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
... [46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
e1-5
1
2010
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
A CD14 gene polymorphism is associated with the IgE level for Dermatophagoides pteronyssinus
2
2005
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Single nucleotide polymorphisms of IL-13 and CD14 genes in allergic rhinitis: a meta-analysis
2
2018
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
... [49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
Association of the CD14 gene polymorphism C-159T with allergic rhinitis
1
2010
... CD14是脂多糖(lipopolysaccharide,LPS)的高亲和力受体,还能够介导其他G+及G-细菌成分与细胞的反应.LPS通过TLR4和cDl4介导刺激IL-12、IFN-γ分泌增加, 可以促使免疫系统向Th1转变.本研究中CD14的rs2563298和rs2569191位点与AR疾病存在阳性相关.尚未有此两位点与AR相关的文献报道,来自埃及[45]和巴基斯坦人群[46]的研究均证实了rs2569191位点与哮喘的相关性,一项爱尔兰的研究[47]证实rs2569191位点与荨麻疹及特异性疾病有关.在本研究中rs2569190位点与AR发病无明显相关,在日本人群研究[48]中结果为阴性,2018年一篇Meta分析[49]同样证实了本观点,但有研究报道不同结果[46,50].一项来自中国北方人群研究[49]发现rs2569190位点TT基因型与AR发病相关,出现矛盾的结果可能由于那项研究的小样本量(病例92例,对照72例)可能导致结果的可信度较低,且中国南北地域差异引起的环境不同. ...
The C-159T polymorphism in the CD14 promoter is associated with serum total IgE concentration in atopic Chinese children
1
2003
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Racial differences in the association of CD14 polymorphisms with serum total IgE levels and allergen skin test reactivity
1
2013
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
A promoter polymorphism in the CD14 gene is associated with elevated levels of soluble CD14 but not with IgE or atopic diseases
1
2004
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
PMID: 23285751
1
2012
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Association of a promoter polymorphism of the CD14 gene and atopy
1
2001
... 在研究TLR2和CD14基因多态性位点与AR相关临床指标的相关性中,我们发现TLR2的rs7656411、CD14的rs2563298和 rs2569191均与AR伴随哮喘的患者有明显相关,证实了诸多TLR2与CD14与哮喘相关的报道[39,44,45,46].在本研究中各多态性位点与总IgE水平无明显相关.在日本[48]、中国儿童[51]、美国[52]以及德国人群[53]研究中同样证实了患者中的血清总IgE与CD14基因多态性无明显相关.然而一些不同人群的研究报道了阳性结果,比如俄罗斯过敏人群[54](TLR2 rs5743708)和美国哮喘人群[55](CD14 rs2569190),这也说明了种族在基因多态性中存在的差异. ...
Toll-like receptor 2 (TLR2) and TLR4 differentially activate human mast cells
1
2003
... 本研究采用MDR模型分析发现TLR2与CD14可能存在基因交互作用.已有多项研究发现TLR2与CD14的协同作用,TLR2是LPS的信号转导分子,它的活化有赖于LPS结合蛋白(LPS binding protein,LBP)和CD14的存在.LPS能够诱导肥大细胞释放 TNF-α、IL-5、IL-10和IL-13,激活TLR2或TLR4信号转导通路可以促进Th2型免疫反应,同时也依赖CD14的存在[56].此外有研究显示过敏性疾病中CD14与环境之间同样有交互作用,皮肤点刺阳性患者中携带CD14 rs2569190 CC基因型且在农村环境可降低哮喘发病率[39].通过MDR模型研究,在菲律宾过敏人群[57](CD14与IL4、FCER1B、 IL4RA、 ADRB2)和韩国过敏儿童人群[58](CD14与IL-4Rα、 IL-13、IL-13Rα1、CTLA4)均发现了CD14与过敏相关基因的交互作用.过敏性疾病本身即是基因与环境共同作用,因此基因与基因、基因与环境交互作用将会更全面阐释过敏疾病发生与发展. ...
Multifactor-dimensionality reduction reveals interaction of important gene variants involved in allergy
1
2015
... 本研究采用MDR模型分析发现TLR2与CD14可能存在基因交互作用.已有多项研究发现TLR2与CD14的协同作用,TLR2是LPS的信号转导分子,它的活化有赖于LPS结合蛋白(LPS binding protein,LBP)和CD14的存在.LPS能够诱导肥大细胞释放 TNF-α、IL-5、IL-10和IL-13,激活TLR2或TLR4信号转导通路可以促进Th2型免疫反应,同时也依赖CD14的存在[56].此外有研究显示过敏性疾病中CD14与环境之间同样有交互作用,皮肤点刺阳性患者中携带CD14 rs2569190 CC基因型且在农村环境可降低哮喘发病率[39].通过MDR模型研究,在菲律宾过敏人群[57](CD14与IL4、FCER1B、 IL4RA、 ADRB2)和韩国过敏儿童人群[58](CD14与IL-4Rα、 IL-13、IL-13Rα1、CTLA4)均发现了CD14与过敏相关基因的交互作用.过敏性疾病本身即是基因与环境共同作用,因此基因与基因、基因与环境交互作用将会更全面阐释过敏疾病发生与发展. ...
Gene-gene interactions between candidate gene polymorphisms are associated with total IgE levels in Korean children with asthma
1
2012
... 本研究采用MDR模型分析发现TLR2与CD14可能存在基因交互作用.已有多项研究发现TLR2与CD14的协同作用,TLR2是LPS的信号转导分子,它的活化有赖于LPS结合蛋白(LPS binding protein,LBP)和CD14的存在.LPS能够诱导肥大细胞释放 TNF-α、IL-5、IL-10和IL-13,激活TLR2或TLR4信号转导通路可以促进Th2型免疫反应,同时也依赖CD14的存在[56].此外有研究显示过敏性疾病中CD14与环境之间同样有交互作用,皮肤点刺阳性患者中携带CD14 rs2569190 CC基因型且在农村环境可降低哮喘发病率[39].通过MDR模型研究,在菲律宾过敏人群[57](CD14与IL4、FCER1B、 IL4RA、 ADRB2)和韩国过敏儿童人群[58](CD14与IL-4Rα、 IL-13、IL-13Rα1、CTLA4)均发现了CD14与过敏相关基因的交互作用.过敏性疾病本身即是基因与环境共同作用,因此基因与基因、基因与环境交互作用将会更全面阐释过敏疾病发生与发展. ...




{kind=link}
{kind=link}